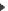 Mardi 8 Mars à 16h, amphi I, Bâtiment 210
Mardi 8 Mars à 16h, amphi I, Bâtiment 210
Sébastien Balibar (LPS-ENS) : Plasticité quantique et supersolidité
Nous avons découvert que, en l’absence totale d’impuretés, les cristaux d’hélium 4 sont anormalement mous. C’est, à notre avis, une conséquence des propriétés quantiques de leurs dislocations, lesquelles peuvent se déplacer par effet tunnel sur des distances macroscopiques (une fraction de millimetre) à grande vitesse (plusieurs mètres par seconde).
Or ce phénomène est lié à une propriété étonnante des cristaux quantiques, leur possible “supersolidité”. Il s’agit cette fois du possible écoulement superfluide d’une partie de la matière à travers le reste du cristal qui reste pourtant élastique, et meme plus rigide que dans son état normal.
D’infimes traces d’impuretés (moins de 1 ppb d’helium 3) suffisent à piéger les dislocations en dessous d’une temperature d’environ 100 millikelvin. Apparement, c’est ce qui permet aux cristaux de passer à l’état “supersolide”, parce que la matière semble couler le long du coeur des dislocations, mais seulement si leur position est fixe.

Un cristal d’hélium 4 vers 100 mK (photo S.Balibar, E. Rolley et C. Guthmann, ENS PAris)
Mardi 18 Janvier à 16h, Amphi II, Bâtiment 210
Hervé Le Treut (IPSL et Ecole Polytechnique) : Effet de serre et évolution climatique future : le diagnostic des modèles
Lundi 15 novembre à 16h, LPT, Bâtiment 210
Frédéric Bournaud (CEA Saclay) : Modéliser les galaxies pour comprendre leur formation
Si le scénario cosmologique standard permet d’expliquer avec succès la formation des grandes structures de l’Univers jusqu’aux amas de galaxies, la formation des galaxies elle-même reste relativement incomprise. Les théories actuelles se révèlent pour le moins infructueuses pour expliquer la morphologie des galaxies, l’histoire cosmologique de leur formation stellaire, et plus globalement la distribution des baryons au sein des
structures de matière noire.
Les propriétés globales des galaxies, telles que leur forme de disque spiral ou de sphéroide elliptique, resultent d’une combinaison complexe de phénomènes à des échelles
très différentes : interactions entre galaxies, courants cosmiques de gaz froid, formation des nuages moléculaires dans les galaxies, formation d’étoiles en amas en leur sein, explosions des supernovae et vents stellaire, sont autant de phénomènes qui jouent tous un rôle majeur dans l’évolution des galaxies. La simulation numérique est un outil indispensable pour comprendre l’intrication de ces processus et tenter d’interpréter les
données observationnelles.
Je présenterai brièvement les techniques principales de simulation numérique dans le domaine, puis, à l’aide de plusieurs exemples résultats récents, montrerai comment les simulations nous permettent de commencer à mieux comprendre la formation des galaxies les plus communes, les galaxies spirales telles que notre propre Voie Lactée. Alors que le paradigme standard consiste à considérer que les propriétés d’une galaxie dépendent
surtout de facteurs extérieurs (interactions et collisions avec d’autres galaxies), les résultats les plus récent montrent que ces propriétés dépendent fortement des propriétés de la turbulence supersonique dans le milieu interstellaire froid et de la physique de la formation stellaire.
Mardi 28 septembre à 16h, LPT, Bâtiment 210
Vincent Croquette (Laboratoire de Physique Statistique, ENS) : Forward and backward motion of replicative polymerases, coupling with their helicase and single molecule version of Sanger sequencing.
In the replisome the polymerase collaborates with the helicase to drive the leading strand synthesis. In vitro, the replicative helicases alone appear to unwind the replication fork with a rate far slower than the normal replicative one and also with a very poor processivity. Replicative polymerases alone are even worse when working on a fork substrate that is in strand displacement. On the other hand, coupling the two enzymes leads to a fast and processive synthesis. The understanding of this coupling is far from perfect and was the motivation of our study.
In this work we investigate replicative polymerases and their helicase coupling in single molecule assays using a DNA hairpin in an unzipping configuration which provides enhance sensitivity. In this simple fork model we assist these enzymes by an external force which provides a control
parameter. If we apply typically 15 pN force on both strands of the DNA hairpin, it opens mechanically. Modulating the force between 0 and 15pN is a mean to assist a molecular motor opening the fork and thus to replace a
partner in the collaborative work. This strategy has already been used to evidence that T7 and T4 helicases display an unwinding rate increasing exponentially with the force. We have conducted this assay with different replicative polymerases, we show that strand displacement polymerization is possible with substantial assisting force level. When the force is reduced the exonuclease activity is activated and the polymerase removes the strand just synthetized.
This finding leads to the development of a Cyclic Polymerase Assay (CPA) where a polymerase is periodically switched from polymerization to strand degradation by modulating the assisting force. Such an assay is very
convenient to study polymerase activity. It allows us to characterize the different polymerase phases (synthesis and exonuclease phases) as a function of force and propose a minimal model for T4 and T7 replicative polymerases.
Moreover, this assay can present very useful applications. In particular we present here a single molecule version of the Sanger sequencing method.
The same assay can also be carried out to study the coupling between helicase and polymerase. We find that the coupled system is very efficient and advances at maximum rate. Coupled replication proceeds at constant rate
without showing any pauses or fork regression, in stark contrast to the case of the isolated polymerase. We explain these results by a collaborative model where both the helicase and the polymerase are described by the
Betterton-Julicher helicase model.




